小麦抗病基因的“贤内助”TdNLR1/TdNLR2 NLR pair协同调控YrTD121位点条锈病抗性
作者: 刁圣轩 审稿人:魏育明 时间: 2025-06-09 点击次数:次
2025年6月9日,四川农业大学刘登才教授团队联合中国科学院遗传与发育生物学研究所刘志勇研究员团队和以色列海法大学Tzion Fahima教授等7家单位合作在Nature Genetics以长文形式发表题为“A head-to-head NLR gene pair from wild emmer confers stripe rust resistance in wheat”的研究论文,揭示了野生二粒小麦中一对呈“头对头”方式排列的新型NLR基因组合TdNLR1和TdNLR2协同调控YrTD121/Yr84位点对条锈病的抗性。其中,TdNLR1为典型的CC-NBS-LRR蛋白,而TdNLR2缺少CC结构域,属于“非典型”NLR;TdNLR1/TdNLR2基因均无与效应因子识别有关的整合结构域。该项工作发现了小麦NLR pair协同调控条锈病抗性新范式,拓展了小麦NLR抗病蛋白抵御病原菌的科学认知。

核苷酸结合富含亮氨酸重复序列(NLR)蛋白是植物中数量最多的免疫受体之一,这类蛋白能够直接或间接地识别病原菌效应子,并激活防御反应。典型的NLR蛋白具有保守的三结构域架构,包含一个N端卷曲螺旋(CC)结构域或Toll-白细胞介素1受体(TIR)结构域、一个中央核苷酸结合(NB-ARC)结构域以及一个C端富含亮氨酸重复序列(LRR)结构域。CC结构域和TIR结构域参与效应子的感知和信号转导;NB-ARC结构域作为NLR激活的分子开关;而LRR结构域在自我抑制中发挥作用,并决定了对效应子识别的特异性。一些NLR蛋白在其结构中还带有额外的整合结构域,这些结构域可充当效应子感受器。
单个NLR蛋白通常能够独立完成对效应子的识别以及下游防御反应的激活。相比之下,某些NLR蛋白可以作为专门的感受器/解码器(Sensor/Executor)NLR pair发挥作用,其中感受器NLR(sensor NLR)识别病原菌效应子,而辅助NLR(helper NLR)则作为则解码器(Executor)诱导下游的免疫反应。大多数已研究的NLR pair在染色体上以“头对头”的方向呈现紧密遗传连锁,并且通过负调控或受体协同作用来发挥功能。在一些情况下,Sensor NLR可以与多个helper NLR蛋白共同发挥作用,从而形成复杂的NLR调控网络。Sensor NLR通常含有整合结构域,例如重金属相关(HMA)结构域、激酶结构域、WRKY结构域、锌指BED结构域以及硝酸盐诱导(NOI)结构域,这些结构域可能来源于受病原菌靶向的宿主蛋白,并作为诱饵识别效应子。
普通小麦(Triticum aestivumL.)提供了人类营养中约20%的热量和蛋白质需求。由条形柄锈菌小麦专化型(Puccinia striiformisWestend. f. sp.tritici,Pst)引起的小麦条锈病对全球小麦生产构成了严重威胁。利用抗病(R)基因培育抗病品种是控制该病害最有效且最环保的策略。目前已正式命名了80多个小麦抗条锈病基因(Yr1-Yr87)。然而,其中只有少数已被克隆,这些克隆的基因编码的要么是单个NLR蛋白,要么是属于不同结构类别的非NLR蛋白。由于病原菌的毒性小种进化迅速,持续努力发掘和克隆更多的抗病基因并将其导入栽培小麦中,对于实现更持久的抗病性至关重要。
野生二粒小麦(T.turgidumssp.dicoccoides)是普通小麦的四倍体野生祖先种,它携带着普通小麦中不存在的新型抗病基因或等位基因。然而,从该物种中分离抗病基因常常面临重组抑制和基因组结构变异的挑战。小麦基因组资源的最新进展以及DNA测序技术的突破,有助于在小麦及其野生近缘种中鉴定新型抗病基因。本研究通过图位克隆和PacBio长读长序列组装相结合的方法,从野生二粒小麦材料TD121(TD104059)中克隆出小麦抗条锈病基因YrTD121。通过EMS诱变、病毒诱导的基因沉默(VIGS)、基于CRISPR/Cas9的基因组编辑以及小麦遗传转化等方法,证明了YrTD121介导的条锈病抗性需要TdNLR1/TdNLR2NLR pair协同作用。YrTD121已被导入栽培小麦中,可用于提高硬粒小麦和/或普通小麦的条锈病抗性。
1.YrTD121赋予小麦对条锈病不完全显性的抗性
野生二粒小麦材料TD121在苗期对条锈菌小种CYR34表现高抗(侵染型(IT)0-9级中,IT为1-2级),且田间成株期对条锈菌混合小种表现免疫(IT为0级)(图1a)。2012-2016年,黄林在以色列海法大学进化所攻读博士学位期间,发现野生二粒小麦TD121携带成株部分抗性基因Yr36,但表现出全生育期的高抗表型。同时,TD121不含已经克隆的全生育期抗病基因Yr15。为了排除Yr36在赋予TD121条锈病抗性中的作用,从TD121与感条锈病的硬粒小麦品种Langdon(LDN)杂交的F5后代中选择出了一个不含Yr36的抗病株系HY-187,并将其与LDN进行回交构建遗传分离群体。虽然HY-187对条锈菌小种CYR34表现高抗(IT为1-2级),但HY-187/LDN的F1仅表现出中抗表型(IT为4-5级,图1a),且F2群体的分离比例为42(抗):66(中抗):37(感病)(χ²₁:₂:₁=0.828,P=0.661),表明该抗性由一个不完全显性基因控制,暂时命名为YrTD121。
2.YrTD121被定位到1BS染色体臂上的1.1 Mb物理区间
为了定位YrTD121,对高抗和高感RNA 混池以及亲本进行了混池分离群体RNA测序(BSR-seq)分析。保留来自每个混池和亲本混池的高质量reads,并将其比对到野生二粒小麦Zavitan参考基因组上。以等位基因频率差异(AFD≥0.7为筛选标准,总共有220个单核苷酸多态性(SNP)位点可能与抗性表型相关;其中130个SNP位点在1B染色体(Chr 1B)上富集,61个聚集在短臂上0至20 Mb的基因组区间内。利用从0至20 Mb基因组区间内鉴定出的SNP或插入缺失(InDel)位点,开发了6个竞争性等位基因特异性PCR(KASP)标记和3个PCR标记,并用于对来自LDN×HY-187的145个F2植株进行基因分型。最近的侧翼标记Sicauw45和Sicauw63将YrTD121定位在3.5 cM的遗传区间内(图1b),对应于Zavitan参考基因组v2.0的3.18 Mb物理区域(1B染色体上9.66–12.84 Mb)。
利用区段标记Sicauw45和Sicauw63对含有5256个单株的F2分离群体进行重组体筛选,获得37个至少代表9种重组类型的重组体。进一步在该区间开发了12个标记,将YrTD121区间缩小到Sicauw76和Sicauw67之间的约1.1 Mb的物理区间(图1c)。
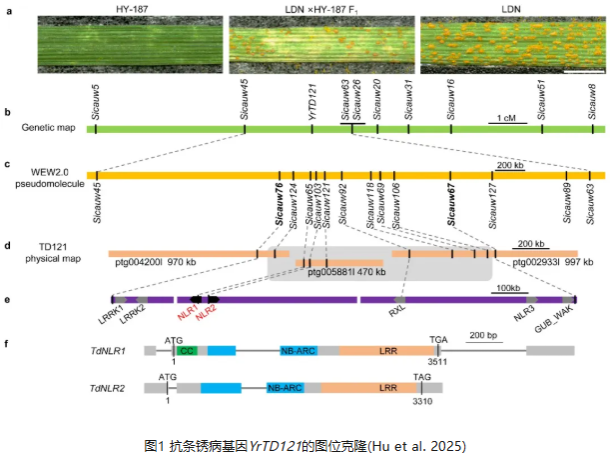
3. PacBio长读长测序和YrTD121的候选基因分析
为了克隆YrTD121,使用PacBioHiFi测序技术对TD121的基因组DNA进行长读长测序。对来自4个SMRT cell(约116.17 Gb)的测序数据组装得到20,653条重叠群(contigs),总组装长度为11.09 Gb,N50重叠群长度为3.06 Mb。使用与YrTD121紧密连锁或共分离的标记作为查询序列,进行本地BLAST搜索,鉴定出三个不重叠的重叠群(ptg004200l,970 kb;ptg005881l,470 kb;以及ptg002933l,997 kb),覆盖YrTD121区域1.18 Mb的基因组序列(图1d)。这些重叠群的序列注释出了7个编码与植物免疫相关蛋白结构域的基因,包括两个富含亮氨酸重复序列的类受体蛋白激酶(LRRK1和LRRK2)、三个NLR蛋白(NLR1、NLR2和NLR3)、一个RX-CC_like(RXL)结构域蛋白,以及一个细胞壁相关受体激酶半乳糖醛酸结合(GUB_WAK_Bind)结构域蛋白(图1e)。
对HY-45、HY-64和HY-187株系的幼苗接种条锈菌小种CYR34后进行RNA测序分析。将得到的reads比对到TD121的PacBio重叠群上,在YrTD121物理区域内鉴定出两个转录的抗病基因,即NLR1和NLR2。这两个NLR基因(此后分别命名为TdNLR1和TdNLR2)以“头对头”的方向排列,被优先作为YrTD121的候选基因进行功能验证(图1e,f)。TdNLR1包含两个外显子,编码一个由961个氨基酸残基组成的典型CC-NB-LRR蛋白,而TdNLR2有三个外显子,编码一个由1002个氨基酸组成的蛋白,具有NB-ARC和LRR结构域。TdNLR1和TdNLR2在蛋白水平上的氨基酸序列一致性约为22%。TdNLR1和TdNLR2都不含有先前报道与病原菌与效应子感知相关的整合结构域。
4.YrTD121候选基因的功能验证
为了研究TdNLR1和TdNLR2在条锈病抗性中的作用,使用甲基磺酸乙酯(EMS)诱变YrTD21在四倍体株系HY-45和六倍体株系绵麦37+YrTD121(MM37+YrTD121),对获得的感病突变体进行TdNLR基因测序。9个突变体(4个四倍体背景,命名为M;5个六倍体背景,命名为EMS)检测到TdNLR1发生点突变且导致编码氨基酸改变(图2a,b);突变体M94和EMS143在TdNLR1编码序列(CDS C1121T,蛋白P374L)中有相同的突变位点。8个突变体在TdNLR2的CDS中有点突变,导致氨基酸改变或翻译提前终止(图2a,c)。这些结果表明,TdNLR1或TdNLR2中的突变都可能导致HY-45和MM37+YrTD121对条锈病抗性的丧失。
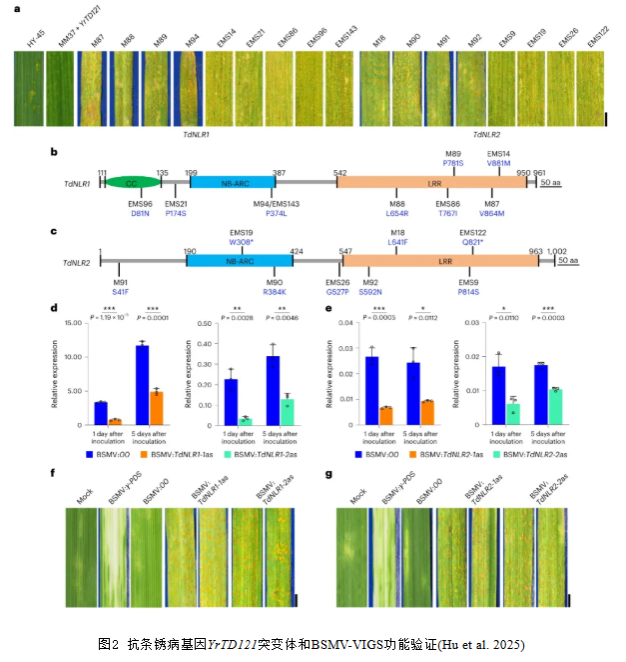
为了进一步验证TdNLR1和TdNLR2在条锈病抗性中的功能,设计了针对TdNLR1和TdNLR2的NB-ARC或LRR结构域的大麦条纹花叶病毒诱导的基因沉默(BSMV-VIGS)载体。用沉默载体接种HY-45导致两个TdNLR基因的转录水平降低了40%-85%(图2d,e)。沉默了TdNLR1或TdNLR2基因的叶片对条锈病的抗性降低,菌孢子堆覆盖的平均叶面积百分比在14.02%到21.14%之间(图2f,g),而阴性对照抗条锈病,表明TdNLR1和TdNLR2是YrTD121介导抗性所必不可少的。
使用CRISPR/Cas9介导的基因组编辑技术在HY-45中对TdNLR1或TdNLR2进行敲除。通过设计了靶向TdNLR基因编码序列的sgRNA,并获得了转基因株系。通过Sanger测序检测靶区域的突变位点。三个TdNLR1突变株系(TdNLR1KO-1到TdNLR1KO-3)分别携带4 bp、7 bp或13 bp的缺失;两个TdNLR2突变株系(TdNLR2KO-1和TdNLR2KO-2)在TdNLR2靶位点分别有一个碱基插入和6 bp缺失。这些基因敲除突变体对接种条锈菌小种CYR34表现为感病,而未编辑的HY-45植株表现为高抗(图3a),这表明TdNLR1和TdNLR2均是HY-45抗条锈病性所必需的。

接下来,使用两个带有自身启动子和下游调控序列的全长TdNLR基因组片段(图3b),以及由玉米泛素启动子驱动的TdNLR1或TdNLR2的开放阅读框(ORF)分别转化感条锈病六倍体小麦品种Fielder。此外,还将带有自身启动子或泛素启动子的TdNLR1和TdNLR2载体共转化到Fielder中。通过PCR鉴定出了单独表达TdNLR1或TdNLR2以及同时表达TdNLR1和TdNLR2的T0代个体。当用条锈菌小种CYR34进行接种时,所有同时表达TdNLR1和TdNLR2或单独表达TdNLR1的T1代植株都具有条锈病抗性,而单独表达TdNLR2的植株则表现为感病(图3c)。在Fielder的1B(TraesFLD1B01G018900)、2B(TraesFLD2B01G357400)和2D(TraesFLD2D01G344800)染色体上分别鉴定出了三个TdNLR2类似的同源基因。1B染色体上的拷贝由于提前终止密码子而被截断。2B和2D染色体上的拷贝与TdNLR2的序列相似性相对较低(约60%),但其预测的蛋白质结构相似,表明它们在赋予抗性方面可能具有与TdNLR2类似的冗余功能。
综上所述,由EMS、VIGS和CRISPR/Cas9介导的突变导致的功能丧失以及转基因互补实验结果表明,TdNLR1对条锈病抗性有正向贡献,而TdNLR2功能的丧失会使YRTD121基因座上TdNLR1介导的抗性丧失。
5.表达分析
使用qRT-PCR测定了YrTD121单基因系HY-187中TdNLR1和TdNLR2的转录水平。TdNLR1和TdNLR2在根、茎和叶中均有表达,在叶片中表达水平最高。在YrTD121纯合子的叶片中检测到的TdNLR1和TdNLR2的表达水平明显高于杂合子。在模拟接种或接种条锈菌后的第1天和第3天,TdNLR1或TdNLR2的表达水平在植株中没有显著差异。
6. TdNLR1和TdNLR2的单倍型和系统发育分析
使用基因特异性引物对906份小麦材料进行筛选,在来自以色列、叙利亚、黎巴嫩和土耳其群体的363份野生二粒小麦材料中,有62份(17%)检测到了TdNLR基因,而在其他小麦亚种或物种的基因型中未检测到。57份野生二粒小麦材料同时含有TdNLR1和TdNLR2基因。使用另外的同源引物扩增测序,发现有两份材料携带TdNLR1和Tdnlr2等位基因;三份材料含有Tdnlr1等位基因和TdNLR2。
对59份野生二粒小麦材料的TdNLR1全长序列进行分析,鉴定出了16种单倍型(TdNLR1_h1到TdNLR1_h16),代表11种推测蛋白单倍型。其中,单倍型TdNLR1_h4、_h5、_h15和_h16在87-107位存在21个核苷酸的缺失,导致与其他单倍型相比,在CC结构域中缺失了7个氨基酸(GVHDDIQ)。对60个TdNLR2序列进行比对,鉴定出了11种单倍型(TdNLR2_h1到TdNLR2_h11),可编码10种蛋白单倍型。TdNLR2_h8在NB-ARC和LRR结构域之间区域有一个‘TA’插入;TdNLR2_h11存在649个核苷酸的缺失,跨越5’非编码区(UTR)的465 bp至起始密码子ATG开始的184 bp编码序列。
11种TdNLR1和10种TdNLR2蛋白单倍型总共代表了20种蛋白单倍型组合(PC1-PC20)。三份材料与TD121蛋白单倍型相同(PC1)。野生二粒小麦材料PI 487260(Yr84)表达TdNLR1和TdNLR2,为单倍型PC2。36份携带不同PC单倍型的材料在田间对条锈病具有抗性。然而,其中一些材料含有如Yr15或Yr36等功能基因。在30份不含Yr15或Yr36的材料中,携带单倍型PC2、PC3、PC4、PC14和PC19的材料对条锈菌具有抗性,表明它们可能是有功能的单倍型。野生二粒小麦WEW110676(PC14)与LDN杂交的F2群体中TdNLR基因型与抗病表型共分离,表明PC14可能是功能单倍型。携带PC7-PC9、PC11、PC13、PC15或PC20的材料表现为感病,表明它们是非功能性单倍型。
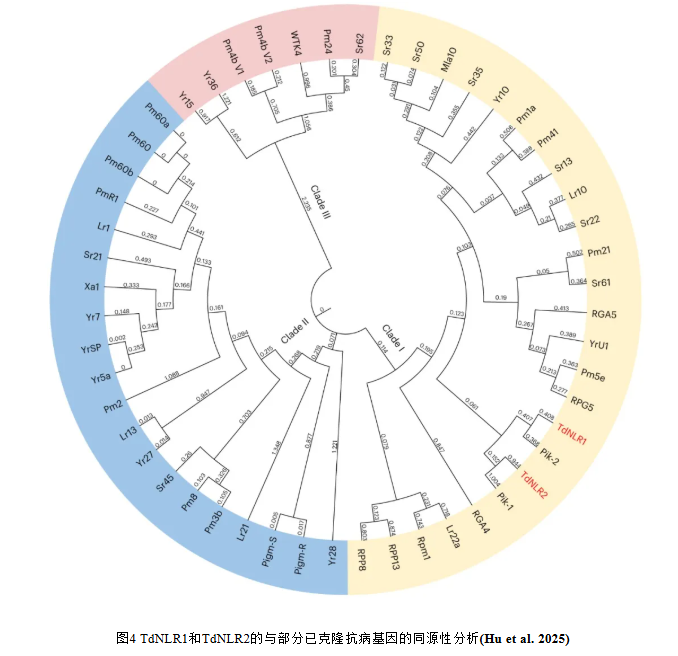
使用TdNLR1和TdNLR2的氨基酸序列作为查询序列,对麦类物种的参考基因组进行BLAST搜索,获得了与TdNLR具有不同相似性水平的同源蛋白。其中,四倍体和六倍体小麦(1B)以及二倍体黑麦(1R)、西尔斯山羊草(Aegilops searsii)、双角山羊草(Ae.bicornis)和高大山羊草(Ae.longissima)(1S)的第1部分同源染色体上的同源蛋白序列与TdNLR1一致性达87.0-90.0%。在2B、2D、5B、5R和5S染色体上发现的其他同源序列与TdNLR1的一致性低于75%。相比之下,与TdNLR2匹配最好的同源序列仅位于1B、1R和1S染色体上,与TdNLR2的一致性低于65%。TdNLR1与高大山羊草中的直系同源蛋白AE.LONG.r1.1SG0000960.1关系最近,而TdNLR2蛋白序列与西尔斯山羊草中的TE01.1S01G0017500.1具有相对较近的进化关系,这表明YRTD121基因座中的TdNLR1和TdNLR2基因可能是从山羊草属物种的1S染色体进化而来的。
为了确定TdNLR pair与禾谷类作物中其他已克隆抗病基因之间的系统发育关系,基于TdNLR1和TdNLR2与50个已发表的抗病蛋白序列的比对构建了系统发育树。鉴定出了三个主要分支(分支I-III),其中分支I和分支II与NLR蛋白聚类在一起,分支III与其他类型的抗病蛋白聚类在一起。TdNLR1和TdNLR2位于分支I中(图4)。与TdNLR1/TdNLR2 pair关系最密切的抗病蛋白是水稻中的Pik-1/Pik-2pair,此前已有报道称其对Pik介导的稻瘟病抗性至关重要。TdNLR1与Pik-2的氨基酸序列一致性为43.3%,而TdNLR2与Pik-1的一致性仅为25.8%,这表明TdNLR1/TdNLR2与Pik-1/Pik/2存在较大的序列分化,但在植物免疫调控中都保留着关键作用。
7. TdNLR1的CC结构域具有诱导细胞死亡的活性且能自我寡聚
为了研究TdNLR1和TdNLR2之间的功能和分子相互作用,在本氏烟草(Nicotiana benthamiana)叶片中瞬时过表达了带有黄色荧光蛋白(YFP)标签的TdNLR全长蛋白以及不同结构域蛋白(图5a)。当YFP标签融合在TdNLR1基因CC结构域序列的C端时,表达TdNLR1_CC-YFP能够诱导细胞死亡(图5b,c);然而,当YFP融合在CC结构域序列的N端时,则不能诱导细胞死亡。有趣的是,表达TdNLR1_h4的CC结构域以及缺失了七个氨基酸(GVHDDIQ)的人工嵌合CC结构域蛋白丧失了诱导细胞死亡的活性。无论是表达TdNLR1的全长、NB、LRR、CC-NB或NB-LRR结构域蛋白,均未观察到细胞死亡现象(图5b)。与TdNLR1的CC结构域不同,在本氏烟草叶片中表达TdNLR2的全长或不同结构域蛋白均未引发坏死反应,这表明其不具有诱导细胞死亡的活性。酵母双杂交和烟草荧光素酶互补分裂实验未能检测到TdNLR1和TdNLR2全长序列之间的直接物理相互作用。
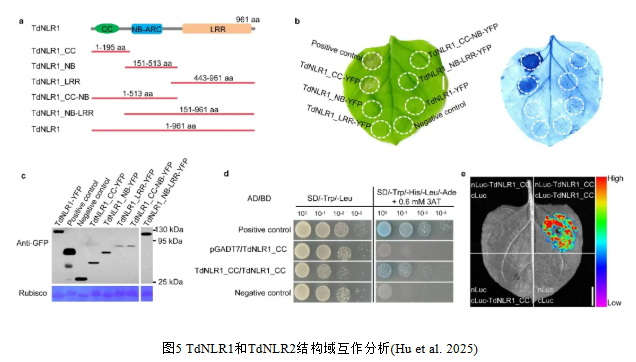
NLR蛋白CC结构域的自身寡聚在触发下游免疫信号中发挥着作用。因此,利用酵母双杂交和荧光素酶互补成像实验来检测TdNLR1的CC结构域是否能自我结合。结果表明,TdNLR1的CC结构域在体外和植物体内能够自我结合(图5d,e)。然而,通过酵母双杂交实验,并未观察到TdNLR1与其CC、NB、LRR、CC-NB和NB-LRR结构域之间的直接相互作用。
8.将YrTD121导入六倍体小麦
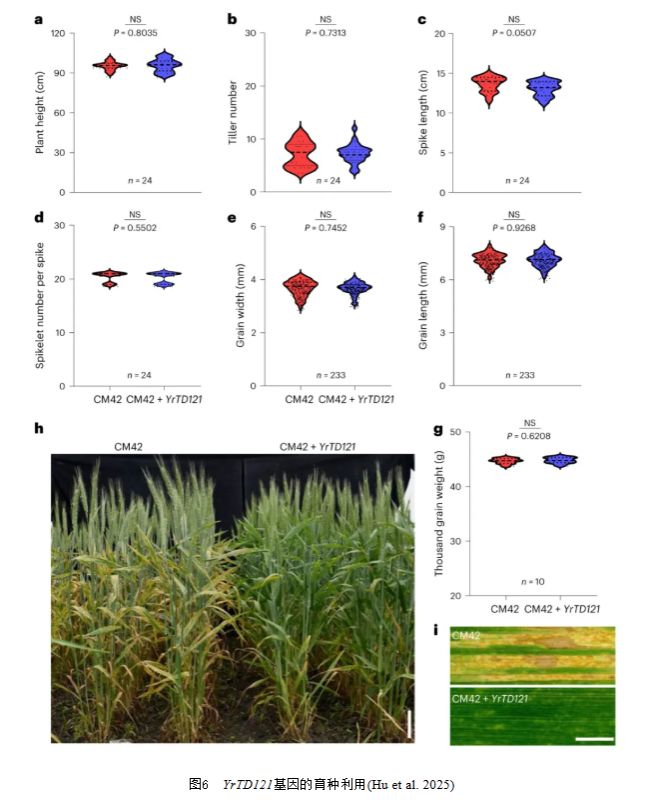
为了便于育种应用,利用侧翼标记Sicauw45和Sicauw26对YrTD121单基因系HY-187与普通小麦品种川麦42(CM42)进行回交筛选。与CM42的中等抗性反应(IT为5-6级)相比,获得的纯合BC3F5株系CM42+YrTD121高抗条锈病(IT为1-2级)(图6)。在调查的七个农艺性状方面,CM42+YrTD121和CM42之间未检测到显著差异(图6)。为了加速YrTD121的育种应用以及与其他抗病基因的聚合,开发了竞争性等位基因特异性PCR(KASP)标记KASP-FM,以便通过分子标记辅助选择来筛选YrTD121。
9.讨论
野生二粒小麦是提高小麦对生物和非生物胁迫耐受性的重要资源。到目前为止,只有少数野生二粒小麦的抗病基因得到了克隆,例如小麦抗条锈病基因Yr15编码的串联激酶WTK1、Yr36基因编码的激酶-START结构域WKS1、Pm36基因编码的串联激酶WTK7-TM,以及编码NLR抗病蛋白的Pm26、Pm41、TdPm60和Pm69。利用长读长测序技术和转录组测序,我们从野生二粒小麦中克隆了抗条锈病基因YrTD121,并证实两个遗传连锁的TdNLR pair是YrTD121介导抗性所必需的。在YRTD121基因座发现的TdNLR pair为小麦抗病育种提供了新抗源。
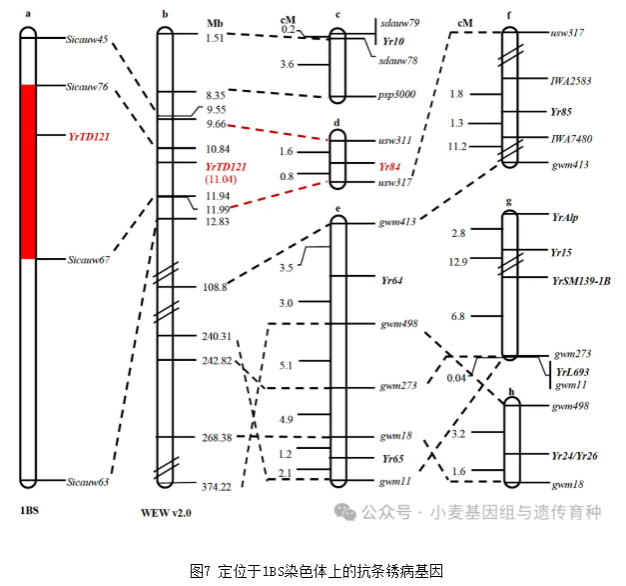
除了在野生二粒小麦材料PI 487260中鉴定出的Yr84,YrTD121的定位区间与大多数先前报道的位于1BS染色体上的抗条锈病基因不同(图7)。对超过300份野生二粒小麦材料进行了筛查,发现PI 487260中存在TdNLR1和TdNLR2基因。在同期背靠背发表的研究论文中,PI 487260中的CNL(TdNLR1)和NL(TdNLR2)pair在赋予YR84基因座的条锈病抗性。对YrTD121和Yr84的NLR pair序列比较显示,TdNLR1存在三个单核苷酸多态性(SNP)位点(A933C、T2184C和A3221G),TdNLR2存在一个SNP位点(A441G)。SNP位点T2184C和A3221G预计会导致TdNLR1中的氨基酸变化(L520S和T866A),SNP位点A441G会导致TdNLR2中出现K110E变异,这意味着YrTD121和Yr84代表不同的功能单倍型。栽培小麦基因库中未检测到YrTD121,表明YrTD121要么未参与栽培小麦驯化,或在小麦驯化过程中丢失。在野生二粒小麦群体中,在YRTD121基因座鉴定出了20种TdNLR蛋白单倍型组合,具有抗性的单倍型PC1(YrTD121)和PC2(Yr84)仅占材料的8%,说明这些基因的频率较低。易感单倍型PC7(TdNLR1_h4 + TdNLR2_h6)占材料的比例高达30.6%。TdNLR1_h4的CC结构域中缺失的七个氨基酸(GVHDDIQ)损害了其诱导细胞死亡的活性,这可能导致了其感病性。在野生二粒小麦中发现的其他潜在功能性单倍型,如PC3、PC4、PC14和PC19,需要进一步的遗传和功能分析。
小麦B亚基因组的直接祖先在449万年前(MYA)与拟斯卑尔脱山羊草(Ae. speltoides)的一个近亲分化开来。在山羊草属的拟斯卑尔脱山羊草组物种中,如高大山羊草(Ae. longissima)和西尔斯山羊草(Ae. searsii),鉴定出了与TdNLR1和TdNLR2最相近的同源基因,这表明单个NLR可能具有古老的起源。然而,YRTD121基因座上的TdNLR1和TdNLR2高度分化,这表明这对TdNLR Pair不太可能是由基因复制产生的。分析发现两份野生二粒小麦材料携带TdNLR1和Tdnlr2等位基因;三份材料携带Tdnlr1等位基因和TdNLR2。有趣的是,TdNLR1和TdNLR2的组合在野生二粒小麦群体中很普遍。因此,“真爱”模型可以解释野生二粒小麦中YRTD121基因座上功能性TdNLR1/TdNLR2 Pair的起源,可能最初它们通过随机的进化事件组合在一起,提供了协同调控的优势,并使它们能够抵御某些病原体。
大多数情况下,单个NLR蛋白感知效应子,随后形成抗病小体以启动细胞死亡。最近的研究表明,一些NLR蛋白可以在遗传连锁的“sensor-helper”Pair或遗传不连锁的“sensor-helper”网络中发挥作用。例如,在水稻和拟南芥中,NLR Pair Pik1/Pik2、CHS3/CSA1、RGA4/RGA5和RRS1/RPS4共同发挥作用。小麦的抗叶锈病基因Lr10需要遗传连锁的RGA2来介导对叶锈病的抗性。本研究中,我们发现野生二粒小麦中以“头对头”方向排列遗传连锁的TdNLR1/TdNLR2Pair介导条锈病抗性。将TdNLR1/TdNLR2随机导入Fielder背景中赋予了条锈病抗性,表明该NLR Pair的头对头排列方向可能对其功能不是那么重要,而TdNLR1/TdNLR2的遗传连锁机制可能有助于该NLR Pair的适应性共进化和遗传共分离,以应对病原菌侵染。单独表达TdNLR1或同时表达TdNLR1和TdNLR2的转基因Fielder植株都具有条锈病抗性,而单独表达TdNLR2的植株则没有抗性,这表明TdNLR2可能作为一个helper NLR与TdNLR1协同发挥作用。先前的报道表明,TNL蛋白SOC3与TN蛋白CHS1或TN2合作,监测E3连接酶SAUL1的稳态;感受器NLR(sensor NLR)蛋白Rx和Bs2可以通过多个helper NLR的旁系同源物(包括NRC2、NRC3或NRC4)互换传递信号。鉴于单独表达TdNLR1的转基因植株具有抗性这一事实,推测Fielder中2B和2D染色体上的TdNLR2旁系同源拷贝可能对TdNLR1介导的免疫具有冗余作用,这需要进一步的研究来解析TdNLR1介导的抗性调控网络。
大多数已研究的NLR Pair包含一个典型的NLR蛋白和另一个带有整合结构域的非典型NLR蛋白,并在植物免疫中作为异源二聚体发挥作用。研究发现,TdNLR1具有典型的NLR结构,而TdNLR2是一个N端截短的NLR蛋白,且没有整合结构域。本研究以及同期背靠背发表的Yr84研究论文,代表了小麦条锈病病理系统中一种没有整合结构域的功能性NLR Pair。尽管TdNLR1和TdNLR2对于赋予YrTD121介导的条锈病抗性都是必需的,但未能检测到它们之间的物理相互作用。在同期背靠背Yr84研究中,检测到了TdNLR1/TdNLR2Pair通过LRR结构域进行不依赖配体的相互作用,这与通过CC或TIR结构域形成异源复合物并相互作用的NLR Pair不同。未来分离出AvrYrTD121效应子并研究它们之间的相互作用,将有助于理解YrTD121的识别和免疫机制。
对导入了YrTD121的品系CM42+YrTD121进行的田间试验初步评估表明,在没有条锈病侵染的情况下,YrTD121对农艺和产量性状没有显著的不利影响,这意味着该基因在小麦育种中具有潜在价值。尽管YrTD121对多种条锈菌小种具有抗性,但新的毒性条锈菌分离株的出现仍有可能使其抗性被克服。将多个抗病基因聚合是维持对植物病害足够且持久抗性的一个有前景的策略。因此,应将YrTD121与具有不同作用方式的慢锈抗性基因,如Yr18、Yr36和Yr46聚合,以实现更持久的抗性。本研究中产生的导入系和开发的分子标记将有助于YrTD121的抗病育种应用。
四川农业大学博士生胡燕灵、中国科学院遗传与发育生物学研究所李淼淼副研究员为论文共同第一作者。四川农业大学小麦研究所黄林副教授、刘登才教授、伍碧华教授、中国科学院遗传与发育生物学研究所刘志勇研究员、以色列海法大学Tzion Fahima教授为论文共同通讯作者。山东农业大学倪飞教授、中国农业科学院作物科学研究所王轲研究员、北京大学现代农业研究院陈时盛研究员、西南科技大学夏崇靖副教授等参与本研究。该研究得到国家自然科学基金(32272068和32341035)、国家重点研发(2023YFD1200402)和西南作物基因资源发掘与利用国家重点实验室等项目资助。
地址: 四川省成都市温江区惠民路211号 四川农业大学小麦研究所 邮编: 611130 电话: 028-82650350