不同田间环境下五个重组自交系群体小麦穗下节间直径的遗传解析及其与农艺性状的关系
作者: 审稿人:刁圣轩 时间: 2021-10-21 点击次数:次
作物产量受源的大小、库的强度、流的流畅度及三者之间的协调性的影响。作物库大、源足,而流不畅会导致空秕率高,进而降低小麦产量。穗下节间是连接旗叶和穗部的桥梁,在小麦源库单位中扮演着流的重要角色。叶片合成及储存在茎中的光合同化物,通过穗下节间运输到穗中,进而促进种子的发育。此外,小麦的茎秆是重要的支撑器官,有利于叶片分布恰当,进而提高光能吸收和光合作用效率。作为从叶片运输光合产物、水分和营养物质到穗部的运输通道,穗下节间对于促进灌浆和提高产量至关重要,是研究源库单位的理想器官。迄今为止,已经报道了许多关于茎秆相关性状位点的研究,然而,很少有研究致力于小麦穗下节间直径的 QTL分析和基因克隆。穗下节间直径的遗传学研究对小麦育种的遗传改良具有重要意义。
本研究利用20828/川农16 (CN16)(2CN, F6) (Ma et al. 2019), 20828/SY95-71 (2SY, F7) (Liu et al. 2020), SHW-L1/川麦32 (CM32)(SC, F8) (Yang 2016; Deng et al. 2019), Q1028/郑麦9023 (ZM9023) (QZ, F18) (Luo et al. 2016), 以及AS985472/苏麦3号 (Sumai 3) (AS, F8) 5个小麦重组自交系群体为材料,对小麦穗下节间直径进行了表型评价(图1),结合5个高密度遗传图谱挖掘与分析控制穗下节间直径的QTL。
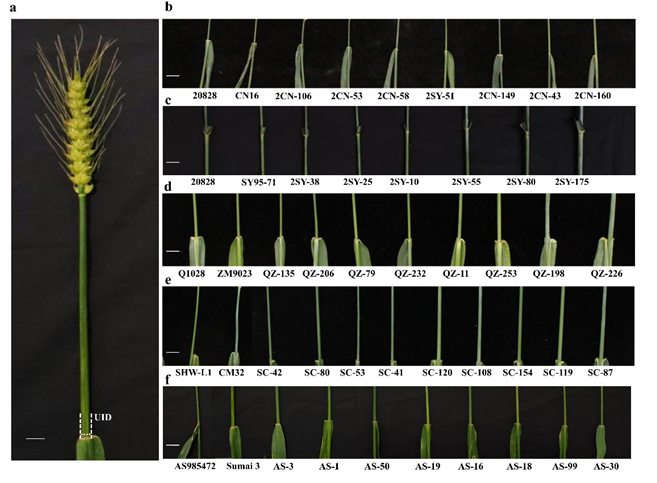
图1 5个重组自交系群体中双亲及部分后代株系的穗下节间直径表型
1. 表型分析
在2CN和2SY重组自交系群体中,20828的穗下节间直径显著大于CN16和SY95-71,2CN群体的穗下节间直径范围为2.32 ~ 4.78 mm,2SY群体的穗下节间直径范围为1.72 ~ 5.18 mm。在QZ群体中,Q1028的穗下节间直径显著高于ZM9023,群体范围为2.10 ~ 5.04 mm。在SC群体中,除了2017CZ,其他环境中的CM32的穗下节间直径均显著高于SHW-L1,群体的穗下节间直径范围为2.14 ~ 5.76 mm。在AS群体中,Sumai 3的穗下节间直径均显著高于AS985472,群体表型范围为2.37 ~ 5.19 mm。2CN, 2SY, SC, QZ以及AS群体的广义遗传力(H2)分别是0.78, 0.69, 0.68, 0.84和0.70,表明5个重组自交系的穗下节间直径主要受遗传因素的影响。5个群体的穗下节间直径表型均表现为正态分布(图2)。
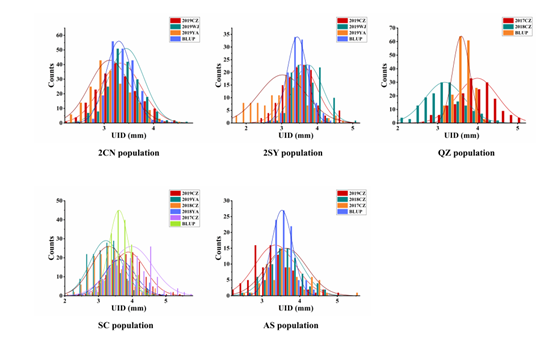
图2 5个重组自交系群体的穗下节间直径频率分布直方图
2. 穗下节间直径和其他农艺性状之间的相关性
相关性结果表明,在2CN群体中,穗下节间直径与穗颈长、株高、开花期、小穗数、穗长、有效分蘖数、旗叶长、和旗叶宽呈极显著正相关性(P < 0.01),在2SY群体中,穗下节间直径与小穗数、旗叶长和旗叶宽呈极显著正相关性,与有效分蘖数呈极显著负相关关系(P < 0.01)。在其他群体中,穗下节间直径与这些农艺性状无显著相关 (P > 0.05)。
3. 穗下节间直径的QTL分析
在5个重组自交系群体中,一共检测到25个控制穗下节间直径的QTL,分布于小麦染色体1A, 1D, 2B, 2D, 3B, 3D, 4A, 4B, 4D, 5A, 5B, 6B和7D上。其中,有5个是主效且稳定表达的穗下节间直径QTL(图3)。
在2CN群体中,检测到13个QTL分布于染色体1A, 1D, 2D, 4A, 4B, 4B, 4D, 5A, 6B和7D上。其中,QUid.sau-2CN-1D.1的等位基因来自于 20828,可在3个环境及BLUP值中检测到,可解释4.43-15.96%的表型变异,LOD值范围为4.76 - 11.92,表明它是一个主效、稳定的QTL。该位点侧翼标记为AX-108775918和AX-111090826。根据QUid.sau-2CN-1D.1的侧翼标记的基因型,将包含199个株系的2CN重组自交系群体分为两个组进行显著性检验。T检验表明,在不同环境及BLUP值中,携带20828等位基因的株系的表型值显著高于携带CN16等位基因的株系的表型值(P < 0.01, Fig. 4-a)。
在2SY群体中,一共定位到2个QTL,分布于1D和5A染色体上。在3个环境及BLUP值中检测到的主效位点QUid.sau-2SY-1D, 可解释13.24 到 30.36% 的表型变异, 该位点定位于标记AX-109130875和AX-111090826之间。QUid.sau-2SY-1D的增效等位基因来源于20828,LOD值范围为4.78 - 9.93。此外,在不同环境及BLUP值中,携带20828等位基因的株系的表型值显著高于携带等位基因的株系的表型值(P < 0.01, Fig. 4-b)。
在QZ群体中,检测到3个QTL:QUid.sau-QZ-2D, QUid.sau-QZ-3B 与 QUid.sau-QZ-5A,可解释5.07 - 14.36%的表型变异。其中,QUid.sau-QZ-2D解释12.13-14.36%的表型变异,LOD值范围为5.31 - 6.89。该QTL的增效等位基因来源于Q1028,定位于标记Xcfd53和Xgpw332之间。在2017CZ 和BLUP值中,该位点携带Q1028等位基因的株系的表型值显著高于携带ZM9023等位基因的株系的表型值(P < 0.01, Fig. 4-c)。
在SC群体中,根据Yu et al. (2015) 的遗传连锁图谱,我们最初将控制穗下节间直径的主效QTL定位到3D染色体短臂上的两个DArT标记之间。由于其中一个侧翼标记的染色体未知,我们进一步利用SC群体3D染色体上的SNP标记 (Yang. 2016) 进行QTL分析。总的来说, SC群体中的79个SNP标记被用于重新构建3D染色体的遗传图谱,该图谱总的遗传距离为 183.31 cM。最终,控制穗下节间直径的主效位点QUid.sau-SC-3D定位于一个0.94 cM的区间内,位于3D染色体短臂上标记AX-109463343和AX-109934976之间。QUid.sau-SC-3D可解释12.86到26.87%的表型变异。其增效等位基因来源于CM32,在6个环境和BLUP值中,QUid.sau-SC-3D携带CM32等位基因的株系的表型值显著高于携带SHW-L1等位基因的株系的表型值(P <0.01,图4-d)。
在AS群体中,一共检测到6个QTL,主效位点QUid.sau-AS-4B解释的表型变异为25.03 - 35.15%。QUid.sau-AS-4B定位于1279649|F|0 - 100007558|F|0,LOD值范围为4.94 - 9.57。该QTL的增效等位基因来源于Sumai 3,携带Sumai 3等位基因的株系表型值显著高于AS985472等位基因的株系表型值 (P < 0.01; Fig. 4-e)。
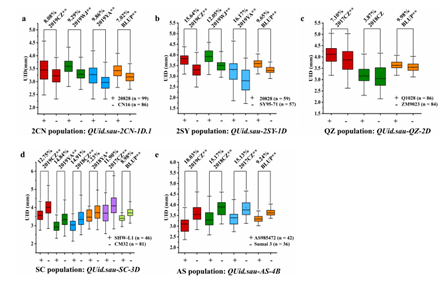
图3 5个重组自交系群体中穗下节间直径主效位点的遗传图谱
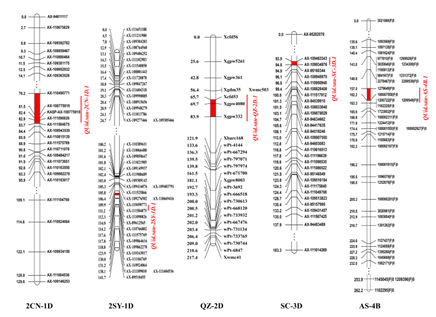
图4 主效QTL的效应分析
4. 主效QTL区间内基因分析
QUid.sau-2CN-1D.1 和 QUid.sau-2SY-1D定位于染色体上相似的物理区间,分别定位于中国春1D长臂上的309.27-316.57 Mb 和316.57-318.74 Mb区间,并且它们的等效等位基因都来源于20828,这两个QTL是同一个位点(QUid.sau-1D)。因此,QUid.sau-1D定位于中国春1D长臂上的309.27-318.74 Mbp和节节麦1D长臂上的314.84 – 324.22 Mbp区间内 (Fig. 5-a)。该位点在中国春区间内有120个候选基因。QUid.sau-QZ-2D定位于标记Xcfd53与Xgpw332之间的一个6.82 cM 的区间,位于中国春2D短臂上的23.02 - 29.84 Mbp和节节麦2D短臂上的24.29 - 497.92 Mb (Fig. 5-b)。由于标记Xgpw332在节节麦上与Xcfd53相隔甚远,其邻近标记Xgpw4080 (在中国春上物理位置为24.49 Mb)被用于确定该QTL的区间。该位点在中国春区间内含有12个候选基因。QUid.sau-SC-3D定位于中国春3D短臂上41.72-43.79 Mb和节节麦3D短臂上41.72-43.79 Mb区间(Fig. 5-c),该位点在中国春区间内有45个候选基因。此外,QUid.sau-AS-4B定位于中国春4B长臂上574.42 - 611.43 Mb 和野生二粒4B 长臂上586.21 - 622.78 Mbp (Fig. 5-d), 中国春区间内注释到380个候选基因。
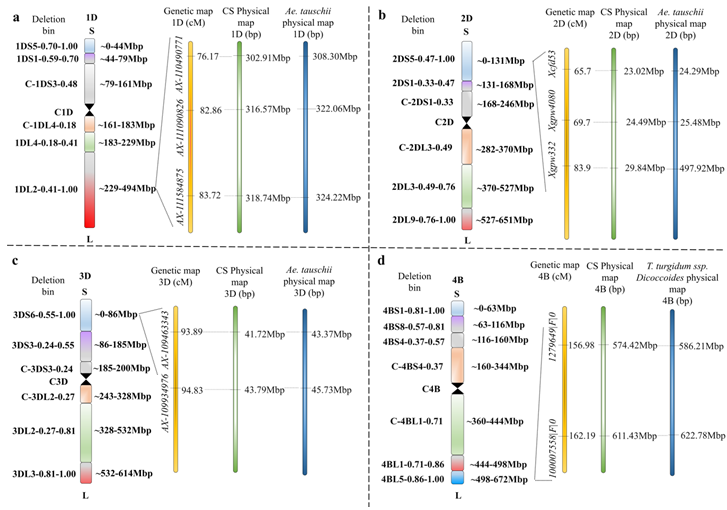
图5 主效QTL的物理图谱
5. 全文总结
本研究分别在5个小麦重组自交系群体中各检测到1个主效且稳定表达的穗下节间直径QTL: QUid.sau-2CN-1D.1, QUid.sau-2SY-1D, QUid.sau-QZ-2D, QUid.sau-SC-3D, 和 QUid.sau-AS-4B。其中QUid.sau-2CN-1D.1 与 QUid.sau-2SY-1D 被证明是同一个 QTL (QUid.sau-1D), 因此这两个QTL可以在两个不同的重组自交系群体中相互验证。我们进一步分析和讨论了小麦穗下节间直径与其他农艺性状之间的关系以及主效QTL区间内与植物生长发育和水分运输有关的候选基因。本研究对控制穗下节间直径位点的挖掘和KASP标记的开发有助于我们了解穗下节间直径的遗传基础,为提高小麦的产量潜力奠定基础。
该结果在Journal of Integrative Agriculture (《农业科学学报》(英文), JIA) 2021年11期正式发表(Genetic dissection of wheat uppermost-internode diameter associated with agronomic traits in five recombination inbred line populations at various field environments)。文章链接:https://www.sciencedirect.com/science/article/pii/S2095311920634128。
该研究得到国家自然科学基金、四川省留学生科技活动重点项目、四川省科技厅应用基础项目、四川农业大学双支计划的资助。
主要参考文献:
Deng M, Wu F Q, Zhou W L, Li J, Shi H R, Wang Z Q, Lin Y, Yang X L, Wei Y M, Zheng Y L,Liu Y X. 2019. Mapping of QTL for total spikelet number per spike on chromosome 2D in wheat using a high-density genetic map. Genetics and Molecular Biology, 42, 603-610.
Goddard M E. 1992. A mixed model for analyses of data on multiple genetic markers.Theoretical and Applied Genetics, 83, 878-886.
Liu J J, Tang H P, Qu X R, Liu H, Li C, Tu Y, Li S Q, Ahsan Habib, Mu Y, Dai S F, Deng M, Jiang Q T, Liu Y X, Chen G Y, Wang J R, Chen G D, Li W, Jiang Y F, Wei Y M, Lan X J, Zheng Y L, Ma J. 2020. A novel, major, and validated QTL for the effective tiller number located on chromosome arm 1BL in bread wheat. Plant Molecular Biology. 104, 173-185
Luo W, Ma J, Zhou X H, Sun M, Kong X C, Wei Y M, Jiang Y F, Qi P F, Jiang Q T, Liu Y X, Peng Y Y, Chen G Y, Zheng Y L, Liu C J, Lan X J. 2016. Identification of quantitative trait loci controlling agronomic traits indicates breeding potential of Tibetan semiwild wheat (Triticum aestivum ssp. tibetanum). Crop Science, 56, 2410-2420.
Ma J, Ding P Y, Liu J J, Li T, Zou Y Y, Habib A, Mu Y, Tang H P, Jiang Q T, Liu Y X, Chen G Y, Wang J R, Deng M, Qi P F, Li W, Pu Z E, Zheng Y L, Wei Y M, Lan X J. 2019. Identification and validation of a major and stably expressed QTL for spikelet number per spike in bread wheat. Theoretical and Applied Genetics, 132, 3155-3167.
Yang J. 2016. QTL mapping for pre-harvest sprouting resistance and molecular characterization of six grain germination-related genes in synthetic wheat. Triticeae Research Institute, Sichuan Agricultural University (Doctoral dissertation,in Chinese).
Yu M, Chen G Y, Pu Z E, Zhang L Q, Liu D C, Lan X J, Wei Y M, Zheng Y L. 2015. Quantitative trait locus mapping for growth duration and its timing components in wheat. Molecular Breeding, 35, 44.
地址: 四川省成都市温江区惠民路211号 四川农业大学小麦研究所 邮编: 611130 电话: 028-82650350