PBJ bHLH转录因子PGS1调控Fl3影响谷物种子大小和重量
作者: 审稿人:刁圣轩 时间: 2022-03-26 点击次数:次
小麦作为三大粮食作物之一,提高其产量对解决粮食短缺问题极为重要。转录因子在谷物发育基因的表达调控中发挥着重要作用。课题组从最初的全基因组小麦转录因子家族分析开始,随后针对其中两个影响种子发育较为突出的转录因子家族进行深入分析,发掘出种子特异表达,潜在的调控种子发育的候选基因,最后进行基因功能验证。
2015年课题组利用小麦表达序列标签(ESTs)共鉴定出2407个可靠的TFs,并进行了系统的分析,将它们划分为63个家族。其中,发育阶段特异性TFs 1257个,组织特异性TFs 1104个。在花、种子、根、叶、茎和根冠中分别鉴定出338、269、262、175、49和18个组织特异性TFs。分别有100、6、342、141、390和278个TFs,在种子休眠、萌发、生殖、成熟、苗期和营养生长阶段特异表达[1]。其中,bHLH(basic helix-loop-helix)家族转录因子参与植物众多生长发育过程如生物与非生物胁迫逆境适应、花发育、种子成熟萌发等调控,该家族成员功能的解析,对作物育种具有巨大潜在价值[2-4]。PLATZ(plant AT-rich zinc-binding proteins)转录因子家族成员较少,是植物中特有转录因子。PLATZ家族序列具有独特的非特异性绑定AT碱基富集区域的特性,同时PLATZ受到ABA调控,也参与植物生长发育调控,如胚乳发育等[5-7]。2017年、2020年课题组利用小麦多组学数据预测了高可信度的225个bHLH[8]和62个PLATZTFs[9],并对其进行特征分析(图1,2)。
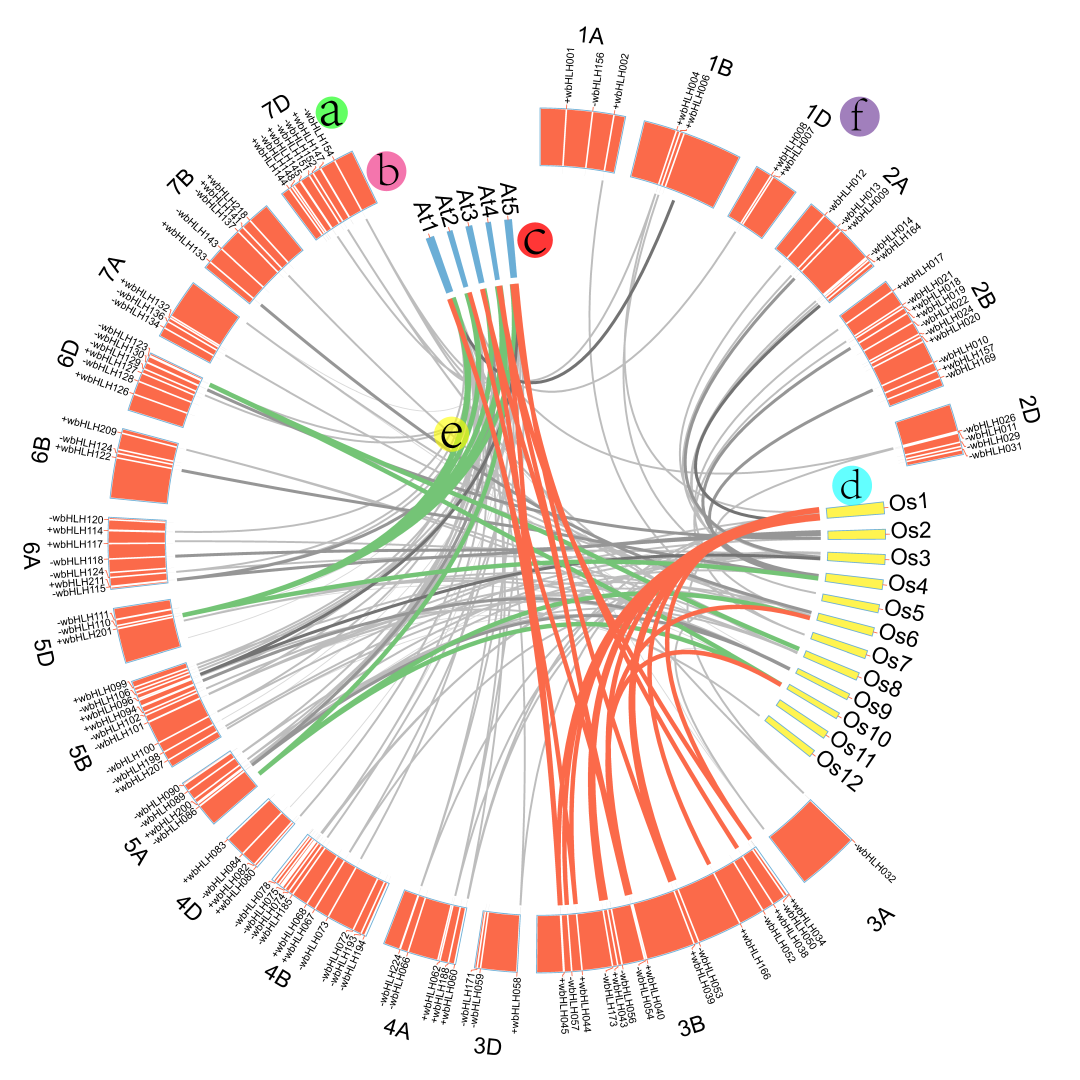
图1.小麦bHLH转录因子在染色体的分布及与水稻、拟南芥的共线性分析[8]。
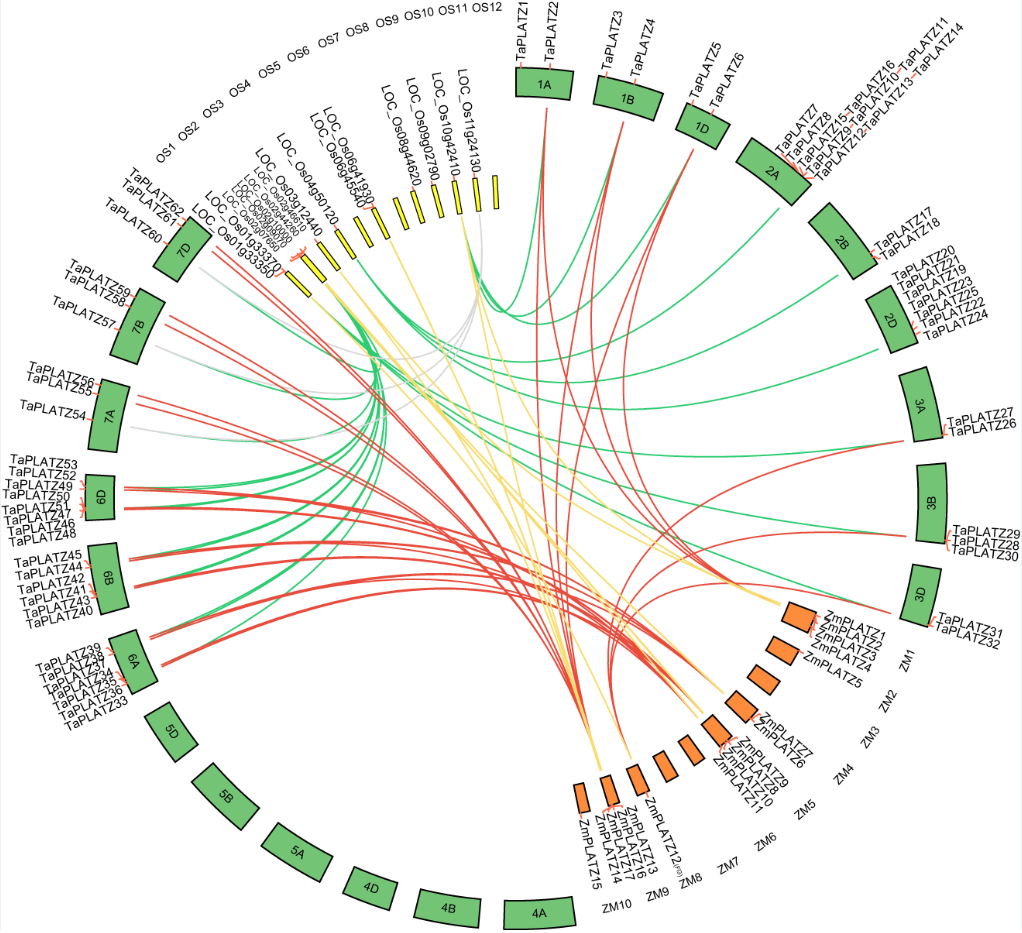
图2.小麦PLATZ转录因子在染色体的分布及与水稻、拟南芥的共线性分析[9]。
2022年3月23日在线发表的论文“The PGS1 basic helix-loop-helix (bHLH) protein regulates Fl3 to impact seed growth and grain yield in cereals”中,报道了TaPGS1在花后5 ~ 20天种子特异性表达,并导致籽粒重量(小麦最高增加13.81%,水稻最高增加18. 55%)和大小增加,碳水化合物和总蛋白质水平也同时增加。胚乳扫描电镜结果表明,TaPGS1过表达小麦和水稻胚乳中淀粉粒较小且紧密嵌在蛋白质之间的间隙中,而株高、穗长、有效穗数和穗粒数等表型在过表达该基因的转基因株系中与野生型相比均无明显差异。TaPGS1直接与小麦、水稻中的PLATZ转录因子TaFl3和OsFl3基因启动子区域的E-box序列结合,积极调控他们在小麦和水稻中的表达,该基因与玉米调控胚乳发育的PLATZ转录因子基因ZmFl3同源。运用CRISPR/Cas9敲除水稻OsFl3,敲除株系平均千粒重、粒宽和粒长均显著下降,而胚乳结构也松散且多空隙,这恰好与TaPGS1的过表达株系表型相反,这进一步证明PGS1通过调控Fl3影响种子的生长发育,进而影响谷物产量(图3,4)。综上,该工作为阐明bHLH和PLATZ转录因子在籽粒发育过程中的潜在功能提供了依据,对提高谷物产量具有重要作用。

图3.过表达TaPGS1增加小麦种子大小和重量[10]。
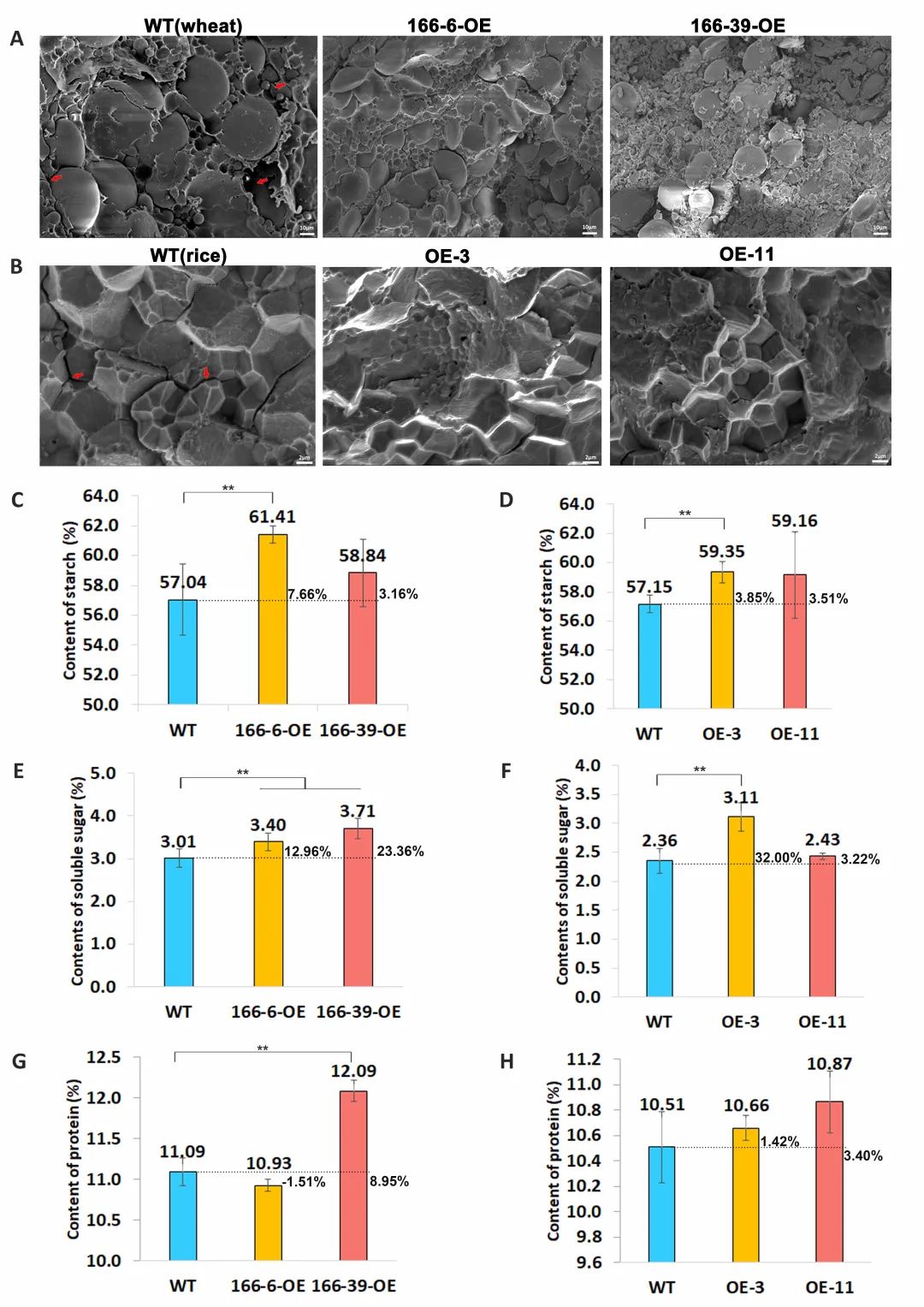
图 4.TaPGS1结合TaFl3和OsFl3启动子促进其表达[10]。
后记:
由于整个课题历时较久,前期受限于小麦基因组较大、测序数据不完整,运用了较老版本的数据库,但是研究思路仍然可取。前期,我们针对性的对bHLH和PLATZ两个家族进行了深入分析。在本研究中,我们结合转录组和ChIPseq分析,确定了PLATZ蛋白Fl3为bHLH转录因子PGS1的下游靶基因。有人可能会问,你们分析了PLATZ家族,为啥靶基因刚好就是这个家族的?这看似是个巧合,却又在情理之中。我们最初选择转录因子家族分析的时候,首先选择的就是与种子发育相关的转录因子家族,而这两个家族均与种子发育调控紧密相关;其次在选择候选基因的时,我们针对性的选择种子特异表达的基因,因此也就造就了这样的“巧合”。
四川农业大学小麦研究所博士生郭晓江、符语昕为论文的共同第一作者,王际睿教授为通讯作者。中国科学院分子植物科学卓越创新中心巫永睿研究员课题组,四川农业大学陈学伟教授课题组,加州大学戴维斯分校柳波教授课题组、加州大学戴维斯分校Mawsheng Chern项目科学家为本研究提供了大力支持。本研究得到了国家重点研发计划“政府间国际科技创新合作/港澳台科技创新合作”重点专项、国家自然科学基金等项目的资助。
原文链接:
https://onlinelibrary.wiley.com/doi/10.1111/pbi.13809?af=R
参考文献:
1. Chen, Z.Y., et al.,Genome-wide characterization of developmental stage- and tissue-specific transcription factors in wheat.BMC Genomics,2015.16: p. 125.
2.Baudry, A., M. Caboche, and L. Lepiniec,TT8 controls its own expression in a feedback regulation involving TTG1 and homologous MYB and bHLH factors, allowing a strong and cell-specific accumulation of flavonoids in Arabidopsis thaliana.Plant J, 2006.46(5): p. 768-79.
3. Grimault A, G.G., et al.,ZmZHOUPI, an endosperm-specific bHLH transcription factor involved in maize seed development.Plant J, 2015.10(1111).
4.Heang, D. and H. Sassa,Antagonistic actions of HLH/bHLH proteins are involved in grain length and weight in rice.PLoS One, 2012.7(2): p. e31325.
5.So, H.-A., et al.,Molecular characterization of stress-inducible PLATZ gene from soybean (Glycine max L.).Plant Omics, 2015.8(6).
6.Nagano, Y., et al.,A novel class of plant-specific zinc-dependent DNA-binding protein that binds to A/T-rich DNA sequences.Nucleic acids research, 2001.29(20): p. 4097-4105.
7.Li, Q., et al.,The Maize Imprinted Gene Floury3 Encodes a PLATZ Protein Required for tRNA and 5S rRNA Transcription through Interaction with RNA Polymerase III.Plant Cell, 2017.29(10): p. 2661-2675.
8.Guo, X.J. and J.R. Wang,Global identification, structural analysis and expression characterization of bHLH transcription factors in wheat.BMC Plant Biol, 2017.17(1): p. 90.
9.Fu, Y., et al.,Identification and Characterization of PLATZ Transcription Factors in Wheat.International Journal of Molecular Sciences, 2020.21(23): p. 8934.
10.Guo, X.J., et al.,The PGS1 basic helix-loop-helix (bHLH) protein regulates Fl3 to impact seed growth and grain yield in cereals. Plant Biotechnology Journal, 2022. doi.org/10.1111/pbj.13809
地址: 四川省成都市温江区惠民路211号 四川农业大学小麦研究所 邮编: 611130 电话: 028-82650350