揭示HvAP2-12/HvAP2-18调控大麦淀粉合成机制
作者: 江千涛 审稿人:魏育明 时间: 2025-09-01 点击次数:次
近日,Plant Biotechnology Journal杂志在线发表了由四川农业大学小麦研究所江千涛教授研究组完成的题为“Dual-Layer Transcriptional-Protein Regulation by HvAP2-12 Represses HvAP2-18 Activity to Fine-Tune Barley Starch Synthesis”的论文。该研究鉴定出一个具有典型转录因子特征的APETALA2/乙烯反应因子(AP2/ERF)家族基因HvAP2-12。过表达HvAP2-12不影响总淀粉和直链淀粉含量,而基因编辑突变株系总淀粉含量增加,直链淀粉含量极显著降低。进一步分析发现,HvAP2-12不直接结合主要淀粉合成基因的启动子,但可结合另一个AP2/ERF家族成员HvAP2-18的启动子,并与HvAP2-18蛋白发生相互作用。HvAP2-18同样影响大麦籽粒淀粉合成,其过表达能提高总淀粉含量,同时降低直链淀粉含量。HvAP2-18直接结合关键淀粉合成基因HvAGP-S1和HvSBE1的启动子,激活其表达。此外,HvAP2-12会抑制HvAP2-18对HvAGP-S1和HvSBE1启动子的结合。基于这些发现, HvAP2-12与HvAP2-18在淀粉基因调控及大麦籽粒品质决定中发挥关键作用。

大麦(Hordeum vulgare L.)是世界第四大作物,广泛用于啤酒酿造和饲料生产。淀粉作为大麦籽粒的主要储存成分,占谷物干重的50%–70%。因此,淀粉含量直接影响籽粒重量,也是构成产量的关键要素。此外,淀粉的组成与结构会影响其理化特性及水解特征,最终决定大麦的营养价值与加工品质。因此,淀粉的含量与特性是决定大麦产量和品质的关键因素。
1. 大麦淀粉调控候选转录因子HvAP2-12的鉴定
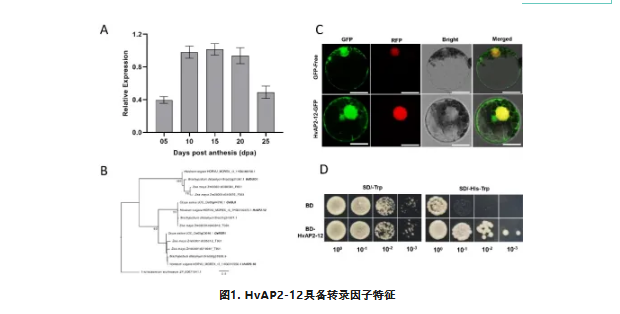
对AP2/ERF转录因子家族进行了全基因组分析,利用已发表的转录组数据,我们筛选出数个在籽粒发育期高表达的AP2/ERF基因。HvAP2-12的表达量在籽粒发育早期(5天)较低,中期(10-20天)达到峰值,后期(25天)逐渐下降(图1A)。氨基酸结构图显示HvAP2-12包含两个AP2/ERF结构域。该基因作为水稻OsGL6的大麦同源基因,可促进水稻叶片毛状体的形成(图1B),但OsGL6在籽粒中的功能尚未被探索,HvAP2-12可能是未被报道参与籽粒淀粉合成的AP2/ERF家族新成员(图1B)。在本氏烟叶片和小麦原生质体中瞬时表达HvAP2-12与绿色荧光蛋白(GFP)的融合蛋白,分析了该蛋白的亚细胞定位。表明HvAP2-12定位于细胞核和细胞质。酵母在筛选培养基上的生长情况及显色反应证实HvAP2-12具有转录激活活性(图1D)。
2. HvAP2-12调控籽粒淀粉与直链淀粉含量
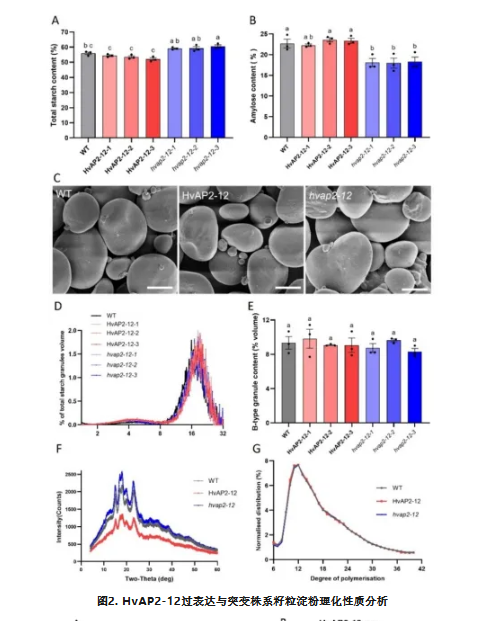
转基因株系与野生型在植株形态、籽粒形态、百粒重及籽粒尺寸(长/宽)方面均未观察到显著差异,表明HvAP2-12不调控这些性状。所有过表达株系的总淀粉含量均降低,而基因编辑株系的总淀粉含量增加(图2A)。与野生型对照相比,HvAP2-12过表达株系的直链淀粉含量未发生显著变化(图2B)。所有突变体株系的直链淀粉含量均极显著下降(图2B)。这些结果表明HvAP2-12显著影响大麦籽粒的淀粉含量与直链淀粉含量。
扫描电镜(SEM)分析显示过表达株系、突变体与野生型具有相似的颗粒结构(图2C)。粒径分布表明HvAP2-12不影响淀粉颗粒形态。X射线衍射(XRD)显示HvAP2-12特异性调控淀粉结晶度而不改变晶体类型。采用高效阴离子交换色谱(HPAEC)对DP6至DP42区间的数据进行标准化处理后发现,过表达株系、突变体与野生型对照之间未出现明显的链长分布模式差异(图2G)。HvAP2-12-2过表达株系、hvap2-12-2突变体与野生型株系无显著区别。
3. HvAP2-12抑制HvAP2-18并与其蛋白互作
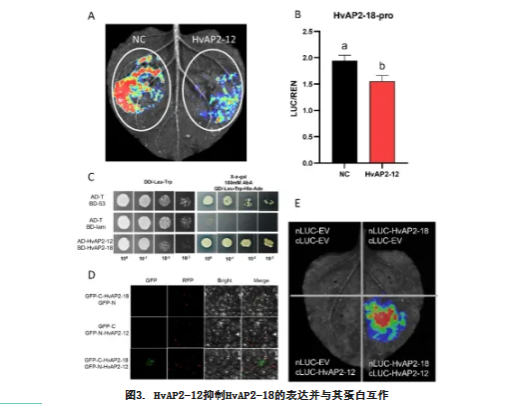
鉴于HvAP2-12不直接结合淀粉合成基因的启动子,我们推测其可能通过调控其他转录因子间接调节这些基因。因此,我们对HvAP2-12转基因转录组数据集中的差异表达基因(DEGs)进行了全面筛选,寻找已知淀粉代谢调控因子(包括AP2/ERF、NAC、MADS和bZIP家族成员)的大麦同源基因。为评估HvAP2-12与这些候选基因的相互作用,我们检测到仅HvAP2-18显示出显著响应表明HvAP2-18直接受HvAP2-12调控(图3A-B)。通过酵母双杂交、双分子荧光互补和荧光素酶互补实验共同证实HvAP2-12与HvAP2-18存在蛋白互作(图3C-E)。
4. HvAP2-18调控淀粉含量与组成但不影响淀粉结构
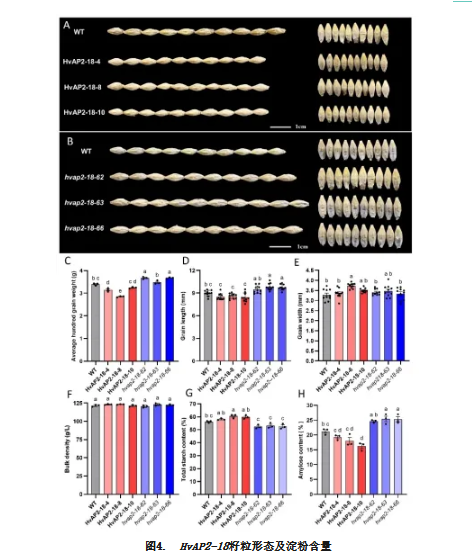
HvAP2-18各株系间籽粒形态存在显著变化(图4A-B)。所有三个突变株系的百粒重均显著增加,而大多数过表达株系的百粒重下降(图4C)。过表达株系的籽粒长度显著减小,而突变株系的籽粒长度较野生型显著增加(图4D)。所有过表达株系或突变株系的籽粒宽度均未呈现一致的显著变化(图4E)。各株系的容重指标无显著差异(图4F)。这些结果表明HvAP2-18在不影响植株整体形态的前提下,能够调控籽粒长度和百粒重。
与野生型相比,大多数HvAP2-18过表达株系的总淀粉含量有所增加(除HvAP2-18-8株系增幅不显著外),而三个突变体的淀粉含量均降低(图4G)。HvAP2-18过表达后直链淀粉含量未呈现一致变化趋势:HvAP2-18-10株系显著低于野生型,但HvAP2-18-4和HvAP2-18-8株系无显著变化。相反,hvap2-18-63和hvap2-18-66突变株系的直链淀粉含量显著增加(图4H)。这些发现表明HvAP2-18同时影响淀粉合成及其组成,但其作用方式与HvAP2-12完全相反。
5. HvAP2-18与淀粉合成基因启动子的结合
通过转录组分析鉴定了HvAP2-18的下游靶基因。表明9个淀粉合成基因在过表达株系中上调而在突变体中下调(图5A),结果证实了HvAP2-18在淀粉合成调控中的作用。双荧光素酶报告基因(DLR)实验证明HvAP2-18能结合HvAGP-S1和HvSBE1启动子并增强其表达(图5B)。启动子截断分析(500 bp×3区段)显示其优先结合HvAGP-S1第一区段和HvSBE1第三区段,该结果通过酵母单杂交实验进一步验证(图5C)。启动子元件分析发现HvAGP-S1第一区段和HvSBE1第三区段均含有STRE元件和GCC-box元件5。我们通过凝胶迁移实验(EMSA)证实HvAP2-18能分别特异性结合HvSBE1启动子中的GCC-box和STRE元件(图5D、E)。虽然HvAP2-12不直接结合这些启动子,但能显著降低HvAP2-18与启动子互作产生的LUC/REN比值(图5F-H),表明HvAP2-12抑制HvAP2-18的调控活性。
6. HvAP2-12通过抑制HvAP2-18协同调控淀粉合成
为研究HvAP2-12与HvAP2-18的相互作用,我们构建了双过表达株系(OE: HvAP2-12-2/HvAP2-18-8)和双突变体(hvap2-12-2/hvap2-18-63)。双过表达株系在粒重、粒长和粒宽方面与野生型无显著差异,而双突变体则表现出百粒重增加和粒长增加。
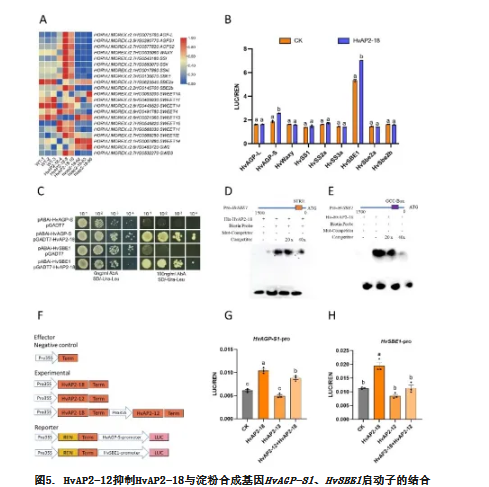
扫描电镜(SEM)分析显示野生型、双过表达株系和双敲除株系间的淀粉颗粒形态无差异(图6A),颗粒粒径分布也保持一致,表明这些基因不影响淀粉结构。淀粉组分分析显示双过表达株系总淀粉含量升高、直链淀粉含量降低(与HvAP2-18过表达株系变化趋势一致;图6B-C),而双突变体总淀粉含量降低、直链淀粉含量升高(与hvap2-18突变体表型相似;图6D-E)。这些结果强有力地证明HvAP2-12在淀粉组成调控中位于HvAP2-18的上游发挥作用。
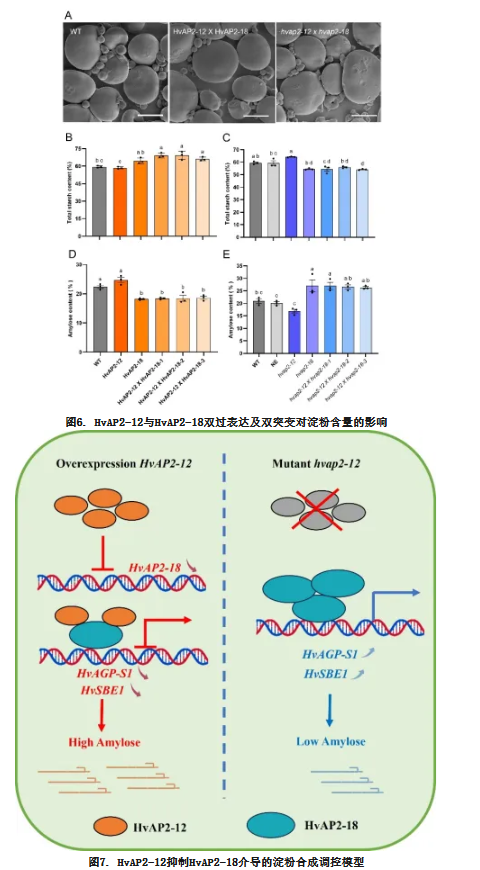
本研究揭示了一种层级抑制调控机制:HvAP2-12通过抑制HvAP2-18的活性间接调控大麦淀粉合成。我们鉴定出这两个AP2家族转录因子是籽粒发育过程中淀粉生物合成的关键调控因子。HvAP2-12通过转录抑制HvAP2-18,而HvAP2-18直接激活淀粉合成关键基因(HvAGP-S1、HvSBE1)的表达。过表达HvAP2-12通过抑制HvAGP-S1/HvSBE1的转录削弱淀粉合成,而hvap2-12基因敲除则增强了HvAP2-18驱动的支链淀粉合成(图7)。
四川农业大学小麦研究所江千涛教授为该文章的通讯作者,已毕业博士研究生丁金金为第一作者。该研究得到了四川省科技支撑计划项目、中国轻工业五粮液浓香型白酒固态发酵重点实验室开放课题的资助。
地址: 四川省成都市温江区惠民路211号 四川农业大学小麦研究所 邮编: 611130 电话: 028-82650350